В качестве интегрирующих элементов, связывающих разные регуляторные механизмы и процесс метаболизма в органах, выступают гормоны. Они исполняют роль химических посредников, обеспечивающих перенос сигналов, возникающих в разных органах и в ЦНС. Клетки по-разному отвечают на действие гормонов. 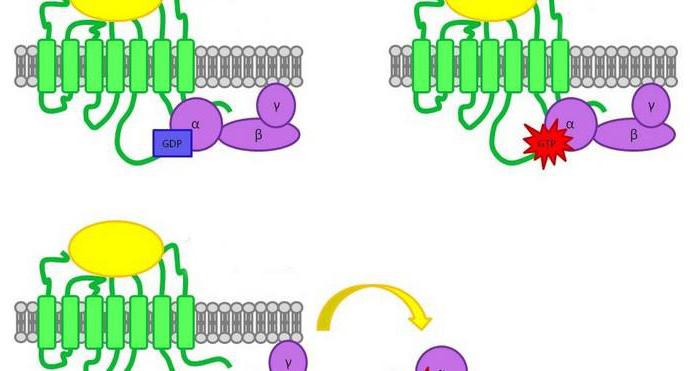
Через аденилатциклазную систему элементы воздействуют на скорость биохимических процессов в клетке мишеней. Рассмотрим эту систему подробно.
Физиологический эффект
Ответная реакция клеток на действие гормонов зависит от его химического строения, а также типа клетки, на которую он воздействует.
Концентрация гормонов в крови достаточно низкая. Для запуска механизма активации фермента с участием аденилатциклазной системы они должны распознаваться, а затем связываться с рецепторами – особыми белками, отличающимися высокой специфичностью.
Физиологический эффект определяется различными факторами, к примеру, концентрацией гормона. Она определяется по скорости его инактивации при распаде, протекающем преимущественно в печени, и скоростью его выведения вместе с метаболитами. Физиологический эффект зависит от степени сродства гормона к белкам-переносчикам. Тиреоидные и стероидные элементы перемещаются по кровеносному руслу совместно с белками. Определяющими факторами являются также количество и тип рецепторов на клетках-мишенях.
Стимулирующие сигналы
Процессы синтеза и секреции гормонов стимулируются внутренними и внешними импульсами, направленными в ЦНС. По нейронам эти сигналы поступают в гипоталамус. Здесь за счет них стимулируется синтез статинов и либеринов (пептидных релизинг-гормонов). Они, в свою очередь, ингибируют (подавляют) либо стимулируют синтез и секрецию элементов в передней доле гипофиза. Эти химические компоненты называются тройными гормонами. Они стимулируют выработку и секрецию элементов в периферических эндокринных железах.
Признаки гормонов
Как и прочие сигнальные молекулы, эти элементы обладают рядом общих черт. Гормоны:
- Выделяются из клеток, их продуцирующих, во внеклеточное пространство.
- Не используются в качестве источника энергии.
- Не являются структурными элементами клеток.
- Обладают способностью устанавливать специфическую взаимосвязь с клетками, имеющими специфические рецепторы для конкретного гормона.
- Отличаются высокой биологической активностью. Даже в незначительных концентрациях гормоны способны эффективно воздействовать на клетки.
Клетки-мишени
Их взаимодействие с гормонами обеспечивается специальными белками-рецепторами. Они находятся на внешней мембране, в цитоплазме, на ядерной мембране и иных органеллах.
В любом белке-рецепторе присутствует два домена (участка). За счет них реализуются функции:
- Узнавания гормона.
- Преобразования и передачи полученного импульса в клетку.
Особенности рецепторов
В одном из доменов белка присутствует участок, комплементарный (взаимодополняющий) какого-то элемента сигнальной молекулы. Связывание рецептора с ней схоже с процессом формирования фермент-субстратного комплекса и определяется показателем константы сродства.
Большая часть рецепторов на сегодняшний день изучена недостаточно. Это связано со сложностью их выделения и очистки, а также крайне низким содержанием в клетках каждого типа рецепторов. Однако известно, что взаимодействие гормонов с рецепторами носит физико-химический характер. между ними образуются гидрофобные и электростатические связи.
Взаимодействие гормона и рецептора сопровождается конформационными изменениями последнего. В результате происходит активация комплекса сигнальной молекулы с рецептором. Находясь в активном состоянии, он способен провоцировать специфический внутриклеточный ответ на поступивший сигнал. При нарушении синтеза или способности рецепторов взаимодействовать с сигнальными молекулами появляются заболевания – нарушения эндокринного характера.
Они могут быть связаны с:
- Недостаточностью синтеза.
- Изменением структуры белков-рецепторов (генетические нарушения).
- Блокированием рецепторов антителами.
Типы взаимодействия
Они различаются в зависимости от строения молекулы гормона. Если она липофильна, то способна проникать сквозь липидный слой в наружной мембране мишеней. Примером являются стероидные гормоны. Если же размеры молекулы значительные, внутрь клетки она проникнуть не может. Соответственно, рецепторы для липофильных гормонов расположены внутри мишеней, а для гидрофильных – снаружи, на внешней мембране.
"Вторые посредники"
Получение ответа на гормональный сигнал от гидрофильных молекул обеспечивается за счет внутриклеточного механизма передачи импульса. Он функционирует за счет так называемых вторых посредников. В отличие от них, молекулы гормонов весьма разнообразны по своей форме.
В качестве "вторых посредников" выступают циклические нуклеотиды (цГМФ и цАМФ), кальмодулин (кальций-связывающий белок), ионы кальция, инозитолтрифосфат, ферменты, принимающие участие в процессе синтеза циклических нуклеотидов и фосфорилирования белков.
Действие гормонов через аденилатциклазную систему
Существует 2 основных способа передачи импульса к клеткам-мишеням от сигнальных элементов:
- Аденилатцеклазная (гуанилатциклазная) система.
- Фосфоинозитидный механизм.
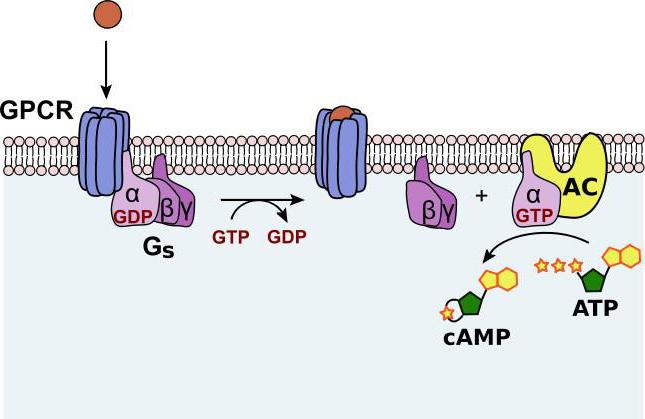
В схеме действия гормонов через аденилатциклазную систему участвуют: G белок, протеинкиназы, белок-рецептор, гуанозинтрифосфат, фермент аденилатцеклаза. Кроме этих веществ, для нормального функционирования системы необходим и АТФ.
Рецептор, G белок, возле которого находятся ГТФ и аденилатциклаза, встроены в клеточную мембрану. Эти элементы находятся в диссоциированном состоянии. После формирования комплекса сигнальной молекулы и белка-рецептора конформация G белка изменяется. В результате одна из его субъединиц приобретает способность взаимодействовать с ГТФ.
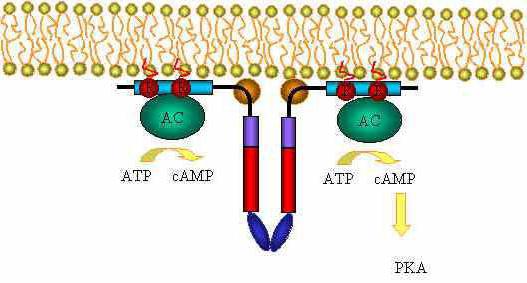
Образовавшийся комплекс "G белок + ГТФ" активирует аденилатциклазу. Она, в свою очередь, начинает трансформировать молекулы АТФ в цАМФ. Она способна активировать специфические ферменты – протеинкиназы. За счет этого катализируются реакции фосфорилирования разных белковых молекул с участием АТФ. В состав белков при этом входят остатки фосфорной к-ты.
За счет механизма действия гормонов в аденилатциклазной системе изменяется активность фосфорилированного белка. В разных типах клеток воздействие оказывается на белки различной функциональной активности: ядерные или мембранные молекула, а также ферменты. В результате фосфорилирования белки могут стать функционально активными либо неактивными.
Аденилатциклазная система: биохимия
За счет взаимодействий, описанных выше, изменяется скорость биохимических процессов в мишени.
Необходимо сказать о незначительной продолжительности активации аденилатциклазной системы. Краткость обусловлена тем, что у G белка после связывания с ферментом начинает проявляться ГТФ-азная активность. Он восстанавливает конформацию после гидролиза ГТФ и перестает воздействовать на аденилатциклазу. Это приводит к прекращению реакции образования цАМФ.
Ингибирование
Кроме непосредственных участников схемы аденилатциклазной системы, в отдельных мишенях присутствуют рецепторы, связанные с G молекулами, приводящие к торможению фермента. Аденилацтеклаза ингибируется комплексом "GTP + G белок".
При остановке выработки цАМФ фосфолирование прекращается не сразу. До тех пор, пока существуют молекулы, будет продолжаться активация протеинкиназ. Для прекращения действия цАМФ клетки используют специальный фермент – фосфодиэстеразу. Он катализирует гидролиз 3',5'-цикло-АМФ до АМФ.
Некоторые соединения, оказывающие угнетающее действие на фосфодиэстеразу (к примеру, теофиллин, кофеин), способствуют сохранению и повышению концентрации цикло-АМФ. Под влиянием этих веществ продолжительность активации аденилатциклазной мессенджерной системы. Другими словами, действие гормона усиливается.
Инозитолтрифосфат
Кроме аденилатциклазной системы трансдукции сигнала, существует еще один механизм передачи сигнала. В нем участвуют ионы кальция и инозитолтрифосфат. Последний представляет собой вещество, производное от инозитфосфатида (сложного липида).
Инозитолтрифосфат образуется под воздействием фосфолипазы "С", специального фермента, который активируется при конформационных изменениях внутриклеточного домена рецептора мембраны клетки.
За счет действия этого фермента гидролизуется фосфоэфирная связь молекулы фосфатидил-инозитол-4,5-бисфосфата. В итоге формируются инозитолтрифосфат и диацилглицерин. Их образование приводит, в свою очередь, к повышению содержания ионизированного кальция в клетке. Это способствует активации различных кальций-зависимых белковых молекул, в числе которых и протеинкиназы.
В этом случае, как и при запуске аденилатциклазной системы, в качестве одного из этапов передачи импульса внутри клетки выступает фосфорилирование белков. Оно приводит к физиологической реакции клетки на воздействие гормона.
Связывающий элемент
В функционировании фосфоинозитидного механизма участвует специальный белок – кальмодулин. Треть его состава образуют отрицательно заряженные аминокислоты (Асп, Глу). В этой связи он способен активно связывать Са+2.
В одной молекуле кальмодулина присутствует 4 связывающих участка. В результате взаимодействия с Са+2 в молекуле кальмодулина начинаются конформационные изменения. В итоге комплекс "Са+2-кальмодулин" приобретает способность регулировать активность многих ферментов: фосфодиэстеразы, аденилатциклазы, Са+2, Mg+2 - АТФазу, а также разные протеинкиназы.
Нюансы
В различных клетках под влиянием комплекса "Са+2-кальмодулин" на изоферменты одного фермента (к примеру, на аденилатциклазу различных типов) в одном случае будет наблюдаться активация, а в другом – угнетение образования цАМФ. Это обуславливается тем, что аллостерические центры в изоферментах могут включать разные радикалы аминокислот. Соответственно, их реакция на воздействие комплекса будет отличаться.
Дополнительно
Как видно, и в аденилатциклазной системе, и в процессах, описанных выше, участвуют "вторые посредники". При функционировании фосфоинозитидного механизма в качестве них выступают:
- Циклические нуклеотиды. Как и в аденилатциклазной системе ими являются ц-ГМФ и ц-АМФ.
- Ионы кальция.
- Комплекс "Са-кальмодулин".
- Диацилглицерин.
- Инозитолтрифосфат. Этот элемент также участвует в передаче сигнала в аденилатциклазной системе.
Механизмы передачи сигналов от молекул гормонов внутри мишеней с участием приведенных выше посредников обладают несколькими общими чертами:
- В качестве одной из стадий передачи информации выступает процесс фосфорилирования белков.
- Активация прекращается под влиянием специальных механизмов. Их запускают сами участники процессов (под воздействием механизмов отрицательной обратной связи).
Заключение
Гормоны выступают в качестве основных гуморальных регуляторов физиологических функций в организме. Они вырабатываются в эндокринных железах или продуцируются специфическими эндокринными клетками. Гормоны выделяются в лимфу, кровь и оказывают дистантное (эндокринное) воздействие на клетки-мишени.
В настоящее время свойства этих молекул достаточно хорошо изучены. Известны процессы их биосинтеза, а также основные механизмы влияния на организм. Однако остается еще немало неразгаданных тайн, связанных с особенностями взаимодействия гормонов и других соединений.