














Родопсин — это распространенный зрительный пигмент, входящий в состав палочковидных зрительных рецепторов сетчатки глаза позвоночных животных. Это вещество имеет очень высокую фоточувствительность и является ключевым компонентом фоторецепции. Иное название родопсина — зрительный пурпур.
В настоящий момент к родопсинам относят пигменты не только палочек, но и рабдомерных зрительных рецепторов членистоногих.
Общая характеристика пигмента
По химической природе родопсин — это мембранный белок животного происхождения, содержащий в своей структуре хромофорную группу. Именно она обуславливает способность пигмента улавливать кванты света. Белок родопсин имеет молекулярную массу примерно 40 кДА и содержит 348 аминокислотных звеньев.
Спектр светопоглощения родопсина состоит из трех полос:
- α (500 нм);
- β (350 нм);
- γ (280 нм).
Лучи γ поглощаются ароматическими аминокислотами в составе полипептидной цепи, а β и α — хромофорной группой.
Родопсин — это вещество, способное распадаться под действием света, что запускает электротонический путь передачи сигнала по нервным волокнам. Данное свойство характерно и для других пигментов фоторецепторов.
Структура родопсина
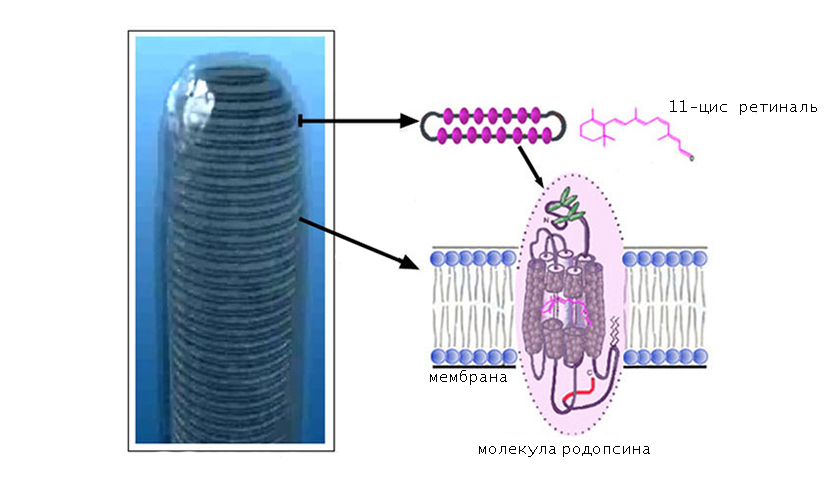
По химической структуре родопсин — это хромогликопротеид, в состав которого входят 3 компонента:
- хромофорная группа;
- 2 олигосахаридные цепочки;
- водонерастворимый белок опсин.
В качестве хромофорной группы выступает альдегид витамина А (ретиналь), который находится в 11-цисформе. Это означает, что длинная часть цепочки ретиналя изогнута и скручена с образованием нестабильной конфигурации.
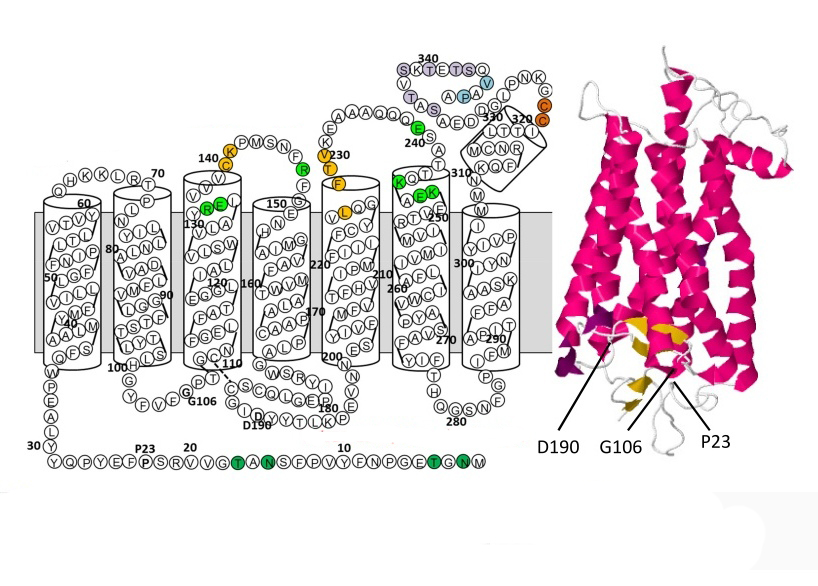
В пространственной организации молекулы родопсина выделяют 3 домена:
- внутримембранный;
- цитоплазматический;
- внутридисковый.
Хромофорная группа расположен во внутримембранном домене. Ее связь с опсином осуществляется через Шиффово основание.
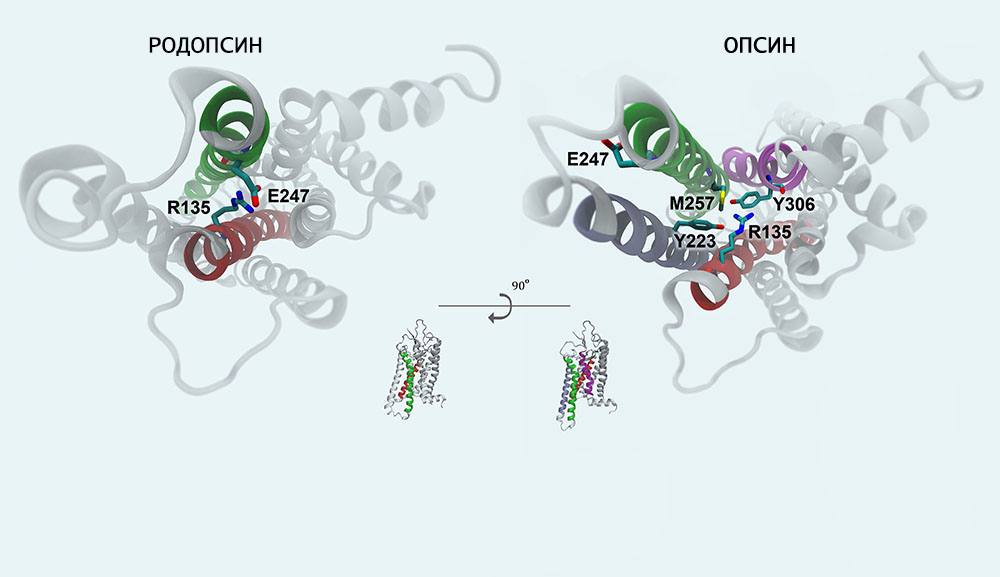
Схема фотопревращения
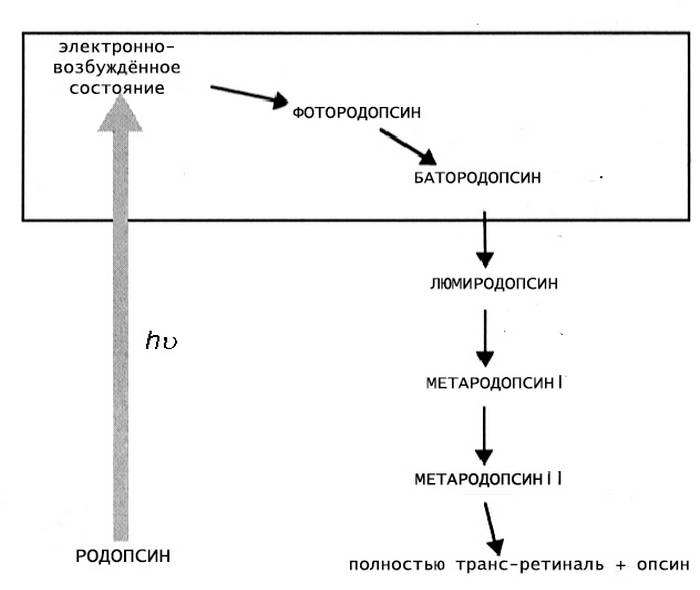
Механизм фотопревращения пигмента родопсина под действием света основан на реакции цис-транс-изомеризации ретиналя — т. е. на конформационном переходе 11-цис-формы хромофорной группы в выпрямленную транс-форму. Этот процесс осуществляется с огромной скоростью (меньше 0,2 пикосекунды) и активирует ряд дальнейших превращений родопсина, которые происходят уже без участия света (темновая фаза).
Образующийся под действием светового кванта продукт называют фотородопсином. Его особенность в том, что транс-ретиналь еще связан с полипептидной цепью опсина.
От завершения первой реакции до конца темновой фазы родопсин последовательно претерпевает следующий ряд превращений:
- фотородопсин;
- батородопсин;
- люмиродопсин;
- метародопсин Ia;
- метародопсин Ib;
- метародопсин II;
- опсин и полностью-транс ретиналь.
Эти превращения сопровождаются стабилизацией, полученной от светового кванта энергии, и конформационной перестройкой белковой части родопсина. В результате хромофорная группа окончательно отделяется от опсина и тут же удаляется из мембраны (транс-форма обладает токсичным действием). После этого запускается процесс регенерации пигмента в исходное состояние.
Регенерация родопсина происходит благодаря тому, что за пределами мембраны транс-ретиналь снова приобретает цис-форму, а затем возвращается обратно, где снова образует с опсином ковалентную связь. У позвоночных восстановление имеет характер ферментативного ресинтеза и происходит с затратой энергии, а у беспозвоночных осуществляется за счет фотоизомеризации.
Механизм передачи сигнала от пигмента в нервную систему
Действующим компонентом запуска фототрансдукции является метародопсин II. В таком состоянии пигмент способен взаимодействовать с белком трансдуцином, тем самым активируя его. В результате связанный с трандуцином ГДФ заменяется на ГТФ. На данной стадии происходит одновременная активизация огромного числа молекул трансдуцина (500—1000). Этот процесс называют первым этапом усиления светового сигнала.
Затем молекулы активированного трансдуцина вступают во взаимодействие с фотодиэстэразой (ФДЭ). Этот фермент в активном состоянии способен очень быстро разрушать соединение цГМФ, необходимое для поддержки в открытом состоянии ионных каналов в мембране рецептора. После вызванной трансдуцином активизации молекул ФДЭ, концентрация цГМФ падает до такого уровня, что каналы закрываются, и в клетку перестают поступать ионы натрия.
Уменьшение концентрации Na+ в цитоплазме наружной части рецептора приводит цитоплазматическую мембрану в состояние гиперполяризации. В результате возникает трансмембранный потенциал, который распространяется до пресинаптического окончания, уменьшая выброс медиатора. Именно это и является смысловым итогом процесса всех преобразований в зрительном рецепторе.











