














Главное условие жизни любого организма - непрерывное поступление энергии, которая расходуется на различные клеточные процессы. При этом определенная часть питательных соединений может использоваться не сразу, а преобразовываться в запасы. Роль такого резервуара выполняют жиры (липиды), состоящие из глицерина и жирных кислот. Последние и используются клеткой в качестве топлива. При этом осуществляется окисление жирных кислот до СО2 и Н2О.
Основные сведения о жирных кислотах
Жирные кислоты представляют собой углеродные цепи различной длины (от 4 до 36 атомов), которых по химической природе относят к карбоновым кислотам. Эти цепи могут быть как разветвленными, так и не разветвленными и содержать разное количество двойных связей. Если последние полностью отсутствуют, жирные кислоты называют насыщенными (характерно для многих липидов животного происхождения), а в противном случае - ненасыщенными. По расположению двойных связей жирные кислоты подразделяют на мононенасыщенные и полиненасыщенные.
Большинство цепей содержит четное число атомов углерода, что связано с особенностью их синтеза. Однако есть соединения с нечетным количеством звеньев. Окисление этих двух типов соединений несколько отличается.
Общая характеристика
Процесс окисления жирных кислот сложный и многостадийный. Он начинается с их проникновения в клетку и завершается в дыхательной цепи. При этом заключительные этапы фактически повторяют катаболизм углеводов (цикл Кребса, превращение энергии трансмембранного градиента в макроэргическую связь). Конечными продуктами процесса являются АТФ, CO2 и вода.
Окисление жирных кислот в клетке эукариот осуществляется в митохондриях (наиболее характерное место локализации), пероксисомах или эндоплазматическом ретикулуме.
Разновидности (типы) окисления
Существует три типа окисления жирных кислот: α, β и ω. Наиболее часто этот процесс протекает по β-механизму и локализуется в митохондриях. Омега-путь представляет собой второстепенную альтернативу β-механизму и осуществляется в эндоплазматическом ретикулуме, а альфа-механизм характерен только для одного вида жирной кислоты (фитановой).
Биохимия окисления жирных кислот в митохондриях
Для удобства процесс митохондриального катаболизма условно подразделяется на 3 этапа:
- активация и транспортировка в митохондрии;
- окисление;
- окисление образовавшегося ацетил-коэнзима А через цикл Кребса и электротранспортную цепь.
Активация представляет собой подготовительный процесс, который переводит жирные кислоты в форму, доступную для биохимических превращений, так как сами по себе эти молекулы инертны. Кроме того, без активации они не могут проникнуть в мембраны митохондрий. Эта стадия протекает у внешней мембраны митохондрий.
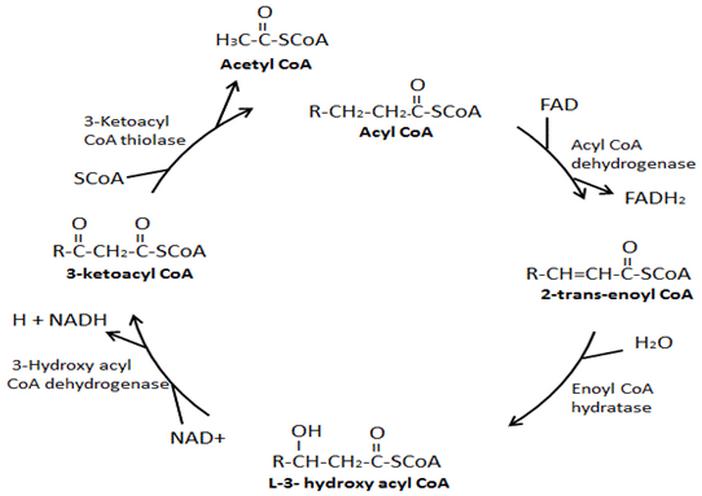
Собственно, окисление - ключевой этап процесса. Оно включает четыре стадии, по окончании которых жирная кислота превращается в молекулы Ацетил-КоА. Тот же продукт образуется и при утилизации углеводов, так что дальнейшие этапы аналогичны последним стадиям аэробного гликолиза. Образование АТФ происходит в цепи переноса электронов, где энергия электрохимического потенциала используется для образования макроэргической связи.
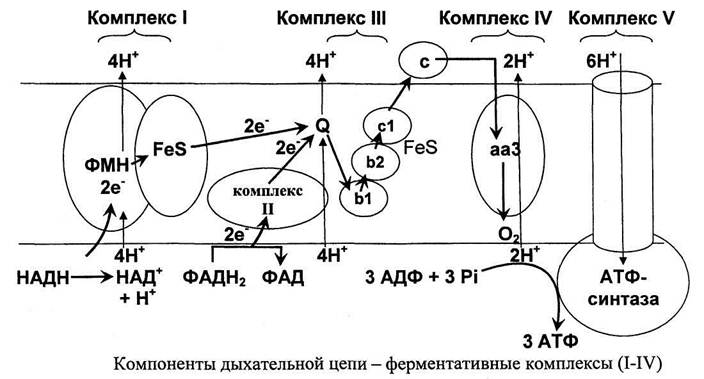
В процессе окисления жирной кислоты кроме Ацетил-КоА образуются также молекулы NADH и FADH2, которые тоже поступают в дыхательную цепь в качестве доноров электронов. В результате суммарный энергетический выход катаболизма липидов достаточно высок. Так, к примеру, окисление пальмитиновой кислоты по β-механизму дает 106 молекул АТФ.
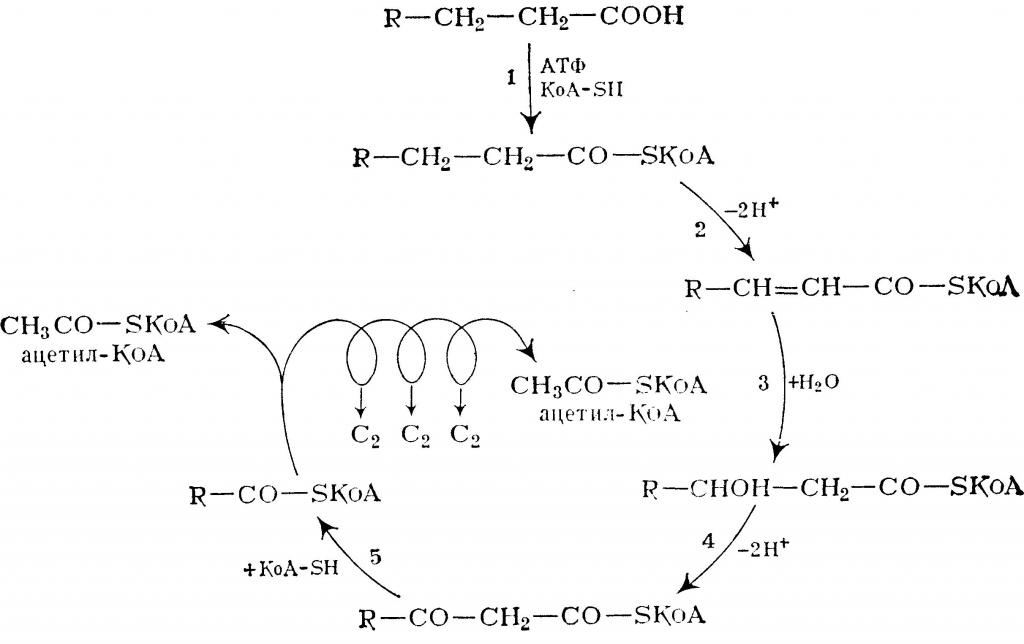
Активация и перенос в митохондриальный матрикс
Жирные кислоты сами по себе инертны и не могут подвергаться окислению. Активация приводит их в форму, доступную для биохимических превращений. Кроме того, в неизменном виде эти молекулы не могут проникнуть в митохондрии.
Суть активации заключается в превращении жирной кислоты в ее Ацил-СоА-тиоэфир, который впоследствии и подвергается окислению. Этот процесс осуществляется специальными ферментами - тиокиназами (Ацил-СоА-синтетазами), прикрепленными к внешней мембране митохондрий. Реакция протекает в 2 этапа, сопряженных с затратой энергии двух АТФ.
Для активации необходимы три компонента:
- АТФ;
- HS-CoA;
- Mg2+.
Вначале жирная кислота взаимодействует с АТФ с образованием ациладенилата (промежуточное соединение). Тот, в свою очередь, реагирует с HS-CoA, тиоловая группа которого вытесняет АМФ, формируя тиоэфирную связь с карбоксильной группой. В результате образуется вещество ацил-CoA - производное жирной кислоты, которое и транспортируется в митохондрии.
Транспортировка в митохондрии
Эта стадия получила название трансэтирификации с карнитином. Перенос ацил-CoA в митихондриальных матрикс осуществляется через поры с участием карнитина и специальных ферментов - карнитин-ацилтрансфераз.
Для транспортировки через мембраны CoA заменяется на карнитин с образованием ацил-карнитина. Это вещество переносится в матрикс методом облегченной диффузии с участием ацил-карнитин/карнитинового переносчика.
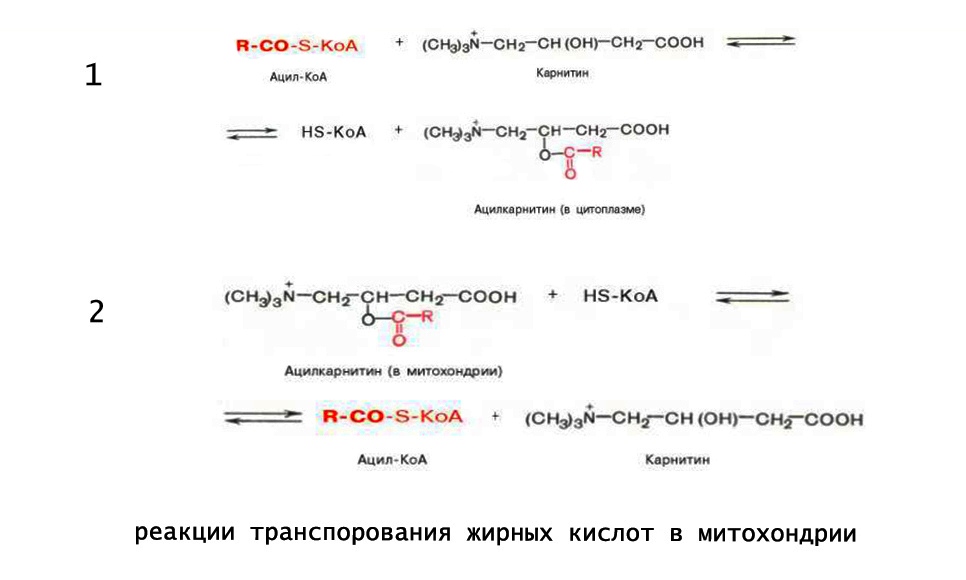
Внутри митохондрий осуществляется реакция обратного характера, заключающаяся в отсоединении ретиналя, вновь поступающего в мембраны, и восстановлении ацил-CoA (в данном случае используется "местный" коэнзим А, а не тот, с которым была образована связь на стадии активации).
Основные реакции окисления жирных кислот по β-механизму
К самому простому типу энергетической утилизации жирных кислот относят β-окисление не имеющих двойных связей цепей, в которых количество углеродных звеньев четное. В качестве субстрата для этого процесса, как уже выше отмечалось, выступает ацил коэнзима А.
Процесс β-окисления жирных кислот состоит из 4 реакций:
- Дегидрирование - отщепление водорода от β-углеродного атома с возникновением двойной связи между звеньями цепи, находящимися в α и β-положениях (первый и второй атомы). В результате образуется еноил-CoA. Ферментом реакции является ацил-CoA-дегидрогеназа, которая действует в комплексе с кофермента ФАД (последний восстанавливается до ФАДН2).
- Гидратация - присоединение молекулы воды к еноил-CoA, в результате чего образуется L-β-гидроксиацил-CoA. Осуществляется еноил-CoA-гидратазой.
- Дегидрирование - окисление продукта предыдущей реакции НАД-зависимой дегидрогеназой с образованием β-кетоацил-коэнзима А. При этом происходит восстановление НАД до НАДН.
- Расщепление β-кетоацил-CoA до ацетил-CoA и укороченного на 2 атома углерода ацил-CoA. Реакция осуществляется под действием тиолазы. Обязательным условием является присутствие свободного HS-CoA.
Затем все снова начинается с первой реакции.
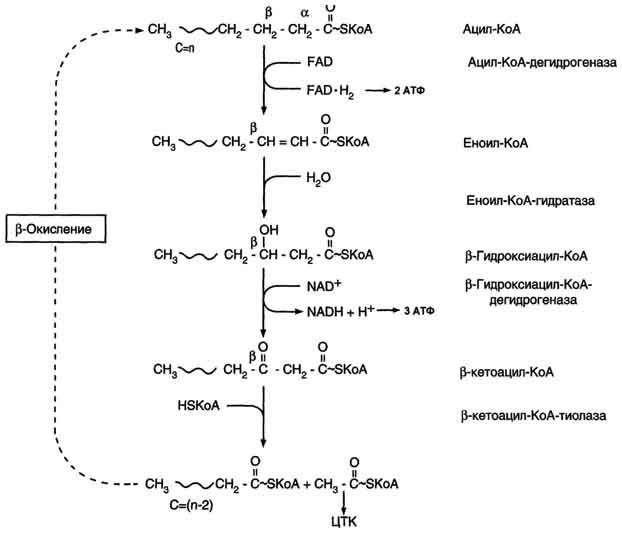
Цикличное повторение всех стадий осуществляется до тех пор, пока вся углеродная цепочка жирной кислоты не превратится в молекулы ацетил-коэнзима А.
Образование Ацетил-КоА и АТФ на примере окисления пальмитоил-CoA
В конце каждого цикла в единственном количестве образуются молекулы ацил-CoA, НАДН и ФАДН2, а цепь ацил-CoA-тиоэфира становится короче на два атома. Передавая электроны в электротранспортную цепь, ФАДН2 дает полторы молекулы АТФ, а НАДН - две. В результате из одного цикла получается 4 молекулы АТФ, не считая энерговыход ацетил-CoA.
В цепочку пальмитиновой кислоты входит 16 углеродных атомов. Это означает, что на стадии окисления должно осуществиться 7 циклов с образованием восьми ацетил-CoA, а энерговыход от НАДН и ФАДН2 в таком случае составит 28 молекул АТФ (4×7). Окисление ацетил-CoA тоже идет на образование энергии, которая запасается в результате поступления в электротранспортную цепь продуктов цикла Кребса.
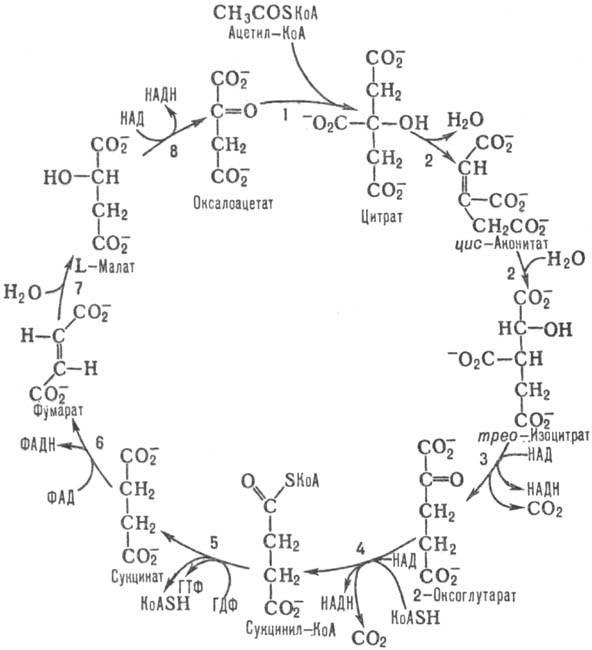
Суммарный выход стадий окисления и цикла Кребса
В результате окисления ацетил-CoA получается 10 молекул АТФ. Так как катаболизм пальмитоил-CoA дает 8 ацетил-CoA, то энергитический выход будет 80 АТФ (10×8). Если сложить это с результатом окисления НАДН и ФАДН2, то получится 108 молекул (80+28). Из этого количества следует вычесть 2 АТФ, которые ушли на активацию жирной кислоты.
Итоговое уравнение реакции окисления пальмитиновой кислоты будет иметь вид: пальмитоил-CoA + 16 О2 + 108 Pi + 80 АДФ = CoA + 108 АТФ + 16 СО2 + 16 H2O.
Расчет выделения энергии
Энергетический выхлоп от катаболизма конкретной жирной кислоты зависит от количества углеродных звеньев в ее цепи. Число молекул АТФ рассчитывается по формуле:
[4(n/2 - 1) + n/2×10] - 2,
где 4 - количество АТФ, образующиеся при каждом цикле за счет НАДН и ФАДН2, (n/2 - 1) - количество циклов, n/2×10 - энерговыход от окисления ацетил-CoA, а 2 - затраты на активацию.
Особенности реакций
Окисление ненасыщенных жирных кислот имеет некоторые особенности. Так, сложность окисления цепей с двойными связями заключается в том, что последние не могут подвергаться воздействию еноил-CoA-гидратазы из-за того, что находятся в цис-положении. Эта проблема устраняется еноил-CoA-изомеразой, благодаря которой связь приобретает транс-конфигурацию. В результате молекула становится полностью идентичной продукту первой стадии бета-окисления и может подвергаться гидратации. Участки, содержащие только одинарные связи, окисляются так же, как насыщенные кислоты.
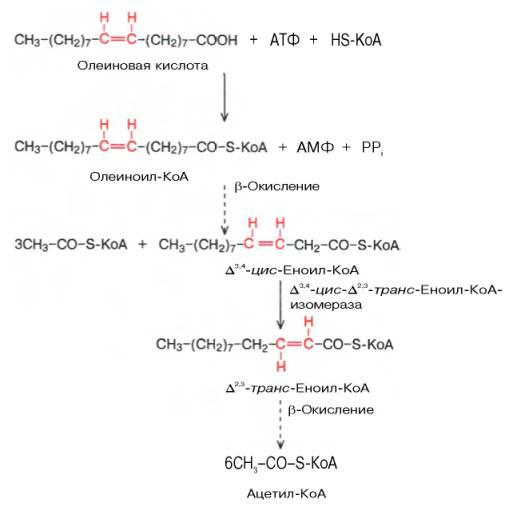
Иногда для продолжения процесса недостаточно еноил-CoA-изомеразы. Это касается цепей, в которых присутствует конфигурация цис9-цис12 (двойные связи при 9-м и 12-м атомах углерода). Здесь помехой является не только конфигурация, но и положение двойных связей в цепи. Последнее исправляется ферментом 2,4-диеноил-CoA-редуктазой.
Катаболизм жирных кислот с нечетным числом атомов
Такой тип кислот характерен для большей части липидов естественного (природного) происхождения. Это создает определенную сложность, так как каждый цикл подразумевает укорачивание на четное число звеньев. По этой причине циклическое окисление высших жирных кислот данной группы продолжается до появления в качестве продукта 5-углеродного соединения, которое расщепляется на ацетил-CoA и пропионил-коэнзим А. Оба соединения поступают в другой цикл из трех реакций, в результате которых образуется сукцинил-CoA. Именно он и поступает в цикл Кребса.
Особенности окисления в пероксисомах
В пероксисомах окисление жирных кислот происходит по бета-механизму, который подобен, но не идентичен митохондриальному. Он также состоит из 4-х стадий, завершающихся образованием продукта в виде ацетил-CoA, но при этом имеет несколько ключевых отличий. Так, водород, отщепившийся на стадии дегидрирования, не восстанавливает ФАД, а переходит на кислород с образованием перикиси водорода. Последний сразу подвергается расщеплению под действием каталазы. В результате энергия, которая могла быть использована для синтеза АТФ в дыхательной цепи, рассеивается в виде тепла.
Второе важное различие заключается в том, что некоторые ферменты пероксисом специфичны к определенным малораспространенным жирным кислотам и отсутствуют в митохондриальном матриксе.
Особенность пероксисом клеток печени заключается в том, что там отсутствует ферментный аппарат цикла Кребса. Поэтому в результате бета-окисления образуются короткоцепочечные продукты, которые для окисления транспортируются в митохондрии.











