














Геном эукариотических клеток помимо информационных последовательностей (экзонов) содержит некодирующие вставки – интроны. Поэтому перед началом белкового синтеза образовавшиеся в результате транскрипции ДНК молекулы подвергаются сплайсингу.
Определение понятия
Сплайсинг – это процесс вырезания из транскрипционной РНК некодирующих участков (интронов) с последующим сшиванием экзонов, что приводит к формированию непрерывной смысловой последовательности, содержащей информацию о первичной структуре белка. Эти манипуляции осуществляются специализированными нуклеопротеидными комплексами – сплайсингосомами.
Сплайсинг – это один из этапов комплексной подготовки рибонуклеиновой кислоты к трансляции, где молекула мРНК служит матрицей, на основе которой в рибосомах по комплементарному принципу строится аминокислотная цепь.
Процессинг РНК
Процессингом называется совокупность пострнанскрипционных модификаций РНК, которые приводят ее к виду, пригодному для участия в белковом синтезе. Иными словами, это процесс превращения первичного транскрипта (пре-мРНК) в матричную РНК. Помимо сплайсинга процессинг включает в себя еще три этапа, к которым относят кэпирование, полиаденелирование и модификацию первичной структуры РНК.
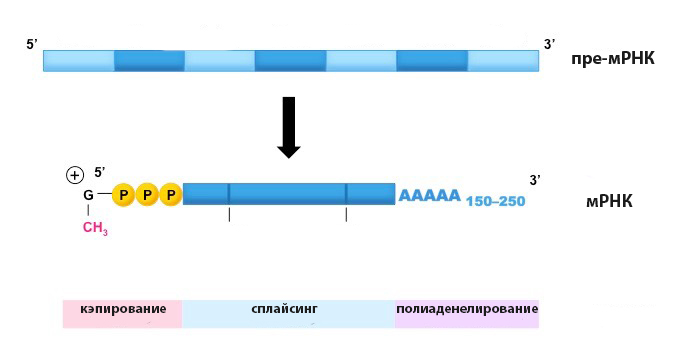
Кэпирование представляет собой образование на 5´-конце РНК особой нуклеотидной последовательности, называемой кэпом (от англ. cap – кепка, шапка). Процесс начинается на этапе транскрипции и осуществляется за счет энергии GTP. Кэп защищает мРНК от нуклеаз, а также выполняет роль сигнального пептида при активации трансляции.
При полиаденелировании фермент поли(А)полимераза присоединяет к 3´-концу РНК остатки адениловой кислоты, в результате чего образуется олиго(А)фрагмент, содержащий от 100 до 250 мономеров (так называемый поли(А)-хвост). Такая конструкция обеспечивает стабильность мРНК в клетке.
Сплайсинг – это третий по очередности процесс, по завершении которого начинается редактирование РНК, включающее модификацию нуклеотидов (метилирование, дезаменирование и т. д.) и вставку внутрь цепи дополнительных азотистых оснований (чаще всего уридиловых). На этом этапе процессинг завершается, и зрелая мРНК выходит из клеточного ядра в цитоплазму.
Сплайсингосомы
В образовании сплайсингосом ключевую роль играют малые ядерные РНК (мяРНК). Из-за большого содержания уридиловых оснований они также называются uPHK (U1,U2, U3 и др.). В комплексе с ядерными белками мяРНК формируют малые рибонуклеопротеиновые частицы (мяРНП), из которых и собираются сплайсингосомы – элипсовидные частицы размером 25 × 5 мкм и коэффициентом седиментации 50-60S.
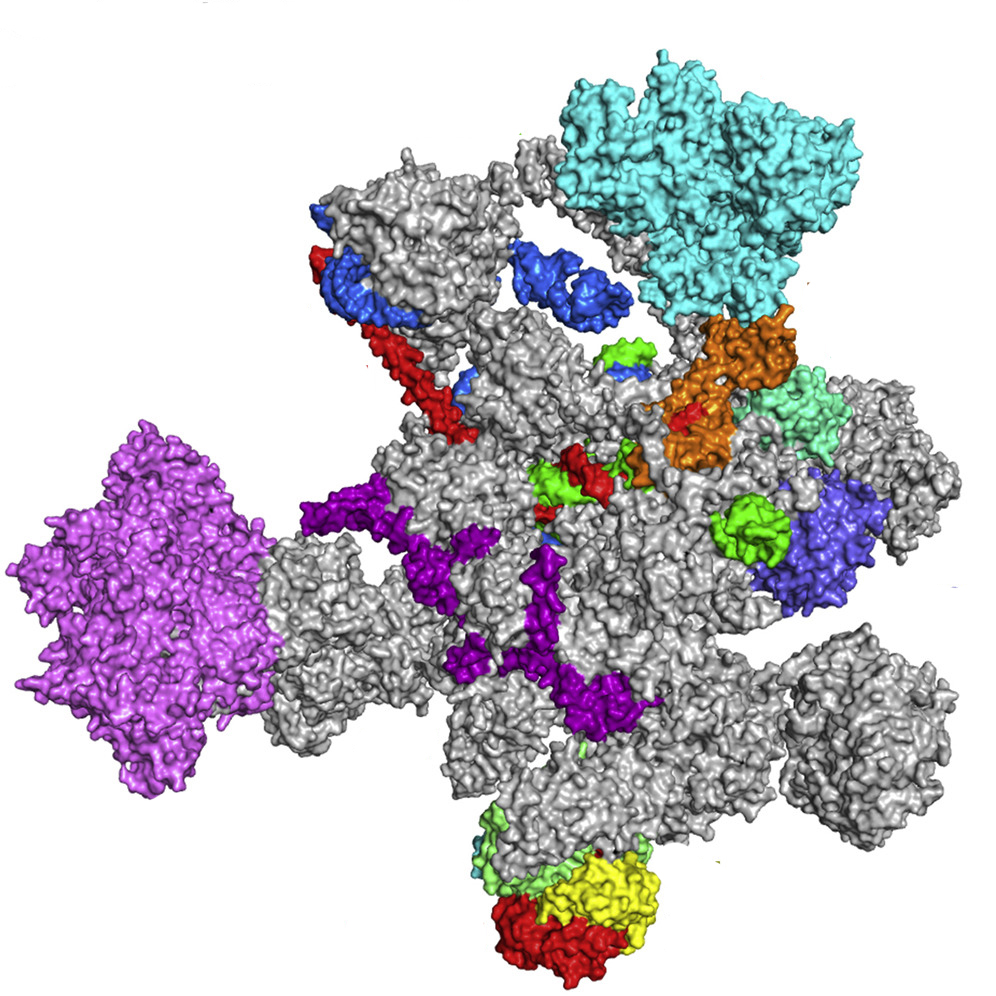
В состав сплайсингосомы млекопитающих входят 6 разновидностей мяРНК (U1–U6). Эти молекулы способны комплементарно взаимодействовать с особыми консервативными последовательностями на концах интронов: GU и AG, называемыми сайтами сплайсинга. Это приводит к выпетливанию и удалению некодирующего участка из матричной последовательности РНК. Сборка нуклеопротеиновых частиц в единый функциональный комплекс происходит непосредственно во время сплайсинга. Стоит отметить, что сама сплайсингосома не разрезает и не сшивает участки РНК, она лишь создает условия для определенного взаимодействия между химическими группами нуклеотидов на концах экзонов и интронов.
Механизм удаления интронов
Возможность сплайсинга РНК во многом определяется особенностью структуры интрона: кроме консенсусных последовательностей (GU на 5´- и AG на 3´-концах) недалеко от 3´-сайта расположен адениловый нуклеотид, входящий в состав сильновырожденной пуриново-пиримидиновой последовательности (PyPyPuAPy...). А-нуклеотид принимает участие в образовании структуры типа "лассо" и называется точкой разветвления. На концах экзонов находятся гуаниновые нуклеотиды, которые в процессе сплайсинга образуют некомплементарную G-G связь.
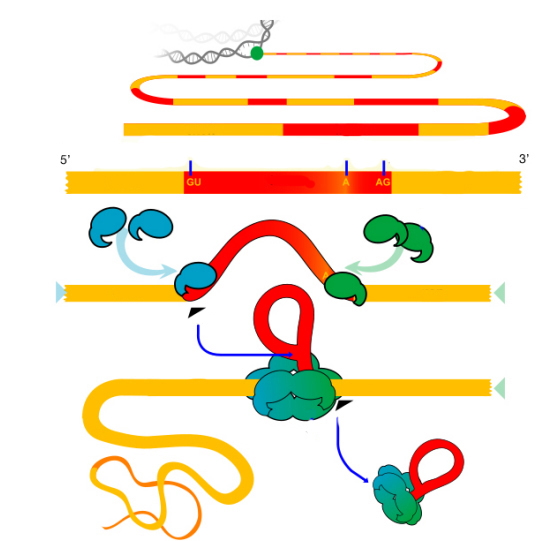
Механизм сплайсинга основан на изменении пространственной конфигурации молекулы РНК. Вначале различные мяРНК комплементарно связываются с GU и AG-сайтами интрона. Параллельно под влиянием структурных белков нуклеопротеидные частицы соединяются в сплайсингосому, из-за чего некодирующий участок РНК дугообазно выгибается, а концы экзонов сближаются с формированием нетипичной водородной связи между гуаниновыми нуклеотидами.
В результате 3´-конец первого экзона оказывается рядом с адениловым нуклеотидом, и фосфодиэфирная связь на границе кодирующей и некодирующей последовательностей разрушается, заменяясь более сильным комплементарным A-G взаимодействием. Таким образом интрон образует петлю в виде лассо. Затем гидроксильная группа освободившегося конца первого экзона атакует 3´-сайт сплайсинга, отщепляя интрон и связываясь со вторым экзоном с образованием цельной РНК.
Альтернативный сплайсинг
Кодирующие участки транскрибированной РНК могут сшиваться в различных комбинациях, формируя альтернативные матричные последовательности. При этом возможно удаление некоторых экзонов или оставление интронов, которые затем участвуют в белковом синтезе. Тип комбинации зависит от выбора сайтов сплайсинга, который регулируется различными белками. Механизм этого процесса в настоящий момент изучен недостаточно. Таким образом, альтернативный сплайсинг представляет собой процесс формирования разных мРНК на основе одного первичного транскрипта.
У эукариот также существуют механизмы формирования альтернативных пре-РНК. К ним относят использование разных промоторов на этапе транскрипции и изменение сайтов полиаденелирования.
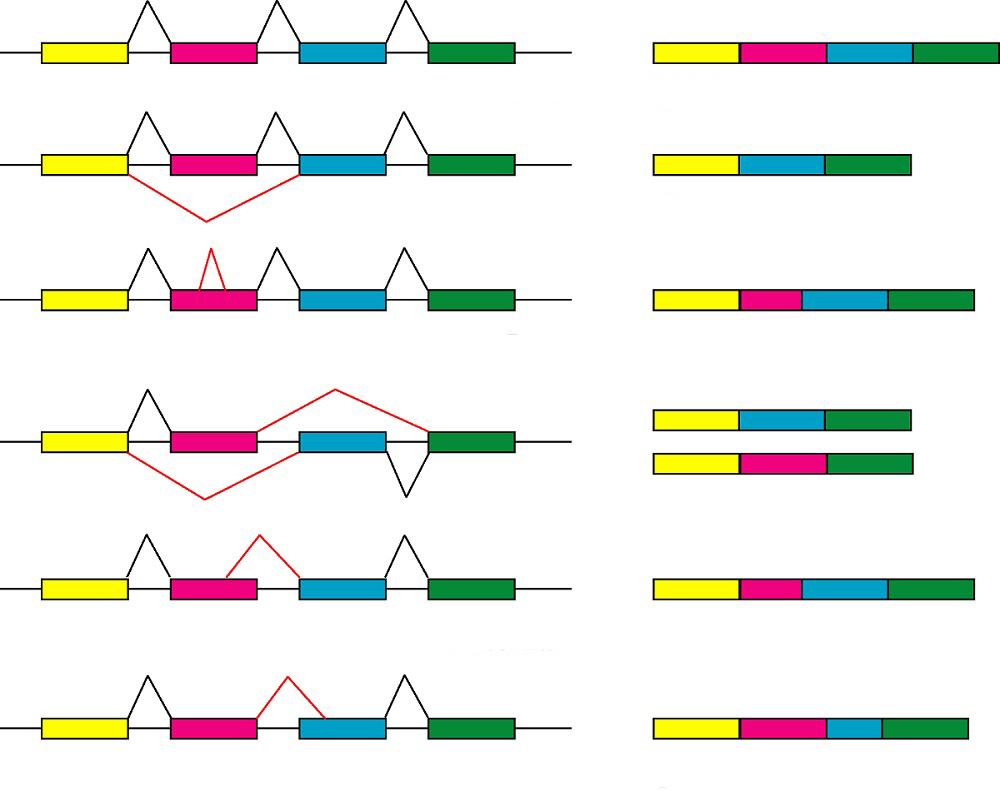
Ключевая роль альтернативного сплайсинга заключается в возможности синтеза нескольких изоформ белка на основе одной смысловой последовательности, что повышает информационную емкость ДНК. Так, у человека благодаря этому механизму 20·103 генов кодируют около 105 типов белков.











