РНК — важнейший компонент молекулярно-генетических механизмов клетки. Содержание рибонуклеиновых кислот составляет несколько процентов ее сухой массы, и около 3-5 % от этого количества приходится на матричную РНК (мРНК), которая непосредственно участвует в белковом синтезе, способствуя реализации генома.
В молекуле мРНК закодирована считанная с гена аминокислотная последовательность белка. Поэтому матричная рибонуклеиновая кислота иначе называется информационной (иРНК).

Общая характеристика
Как и все рибонуклеиновые кислоты, матричная РНК представляет собой цепь рибонуклеотидов (аденин, гуанин, цитозин и урацил), соединенных друг с другом фосфодиэфирными связями. Чаще всего мРНК имеет только первичную структуру, но в некоторых случаях — вторичную.

В клетке присутствуют десятки тысяч видов иРНК, каждый из которых представлен 10-15 молекулами, соответствующими конкретному участку в ДНК. В иРНК записана информация о структуре одного или нескольких (у бактерий) белков. Аминокислотная последовательность представлена в виде триплетов кодирующего участка молекулы мРНК.
Биологическая роль
Основная функция матричной РНК заключается в реализации генетической информации путем ее передачи от ДНК к месту синтеза белка. При этом иРНК выполняет две задачи:
- переписывает информацию о первичной структуре белка с генома, что осуществляется в процессе транскрипции;
- взаимодействует с белковосинтезирующим аппаратом (рибосомами) в качестве смысловой матрицы, определяющей последовательность аминокислот.
Собственно, транскрипция представляет собой синтез РНК, в котором ДНК выступает в качестве матрицы. Однако только в случае информационной РНК этот процесс имеет значение переписывания с гена информации о белке.
Именно иРНК является главным посредником, при помощи которого осуществляется путь от генотипа к фенотипу (ДНК-РНК-белок).

Время существования мРНК в клетке
Матричная РНК живет в клетке очень недолго. Период существования одной молекулы характеризуется двумя параметрами:
- Функциональное время полужизни определяется способностью мРНК служить в качестве матрицы и измеряется по уменьшению количества белка, синтезируемого с одной молекулы. У прокариот этот показатель составляет примерно 2 минуты. За этот период количество синтезируемого белка уменьшается в два раза.
- Химическое время полужизни определяется уменьшением молекул информационных РНК, способных к гибридизации (комплементарному соединению) с ДНК, что характеризует целостность первичной структуры.
Химическое время полужизни обычно длиннее функционального, поскольку незначительная начальная деградация молекулы (например, единичный разрыв в рибонуклеотидной цепи) еще не препятствует гибридизации с ДНК, но уже препятствует белковому синтезу.
Время полужизни является статистическим понятием, поэтому существование конкретной молекулы РНК может оказаться значительно выше или ниже этого значения. В результате некоторые мРНК успевают транслироваться несколько раз, а другие деградируют раньше, чем закончится синтез одной белковой молекулы.
В плане деградации эукариотические иРНК гораздо стабильнее прокариотических (период полужизни составляет около 6 часов). По этой причине их гораздо легче выделять из клетки в неповрежденном виде.
Структура мРНК
В нуклеотидную последовательность матричной РНК входят транслируемые области, в которой закодирована первичная структура белка, и неинформативные участки, состав которых у прокариот и эукариот отличается.
Кодирующая область начинается с инициирующего кодона (AUG) и заканчивается одним из терминирующих (UAG, UGA, UAA). В зависимости от типа клетки (ядерная или прокариотическая) матричная РНК может содержать один или несколько транслирующих участков. В первом случае она называется моноцистронной, а во втором - полицистронной. Последняя характерна только для бактерий и архей.
Особенности строения и функционирования мРНК у прокариот
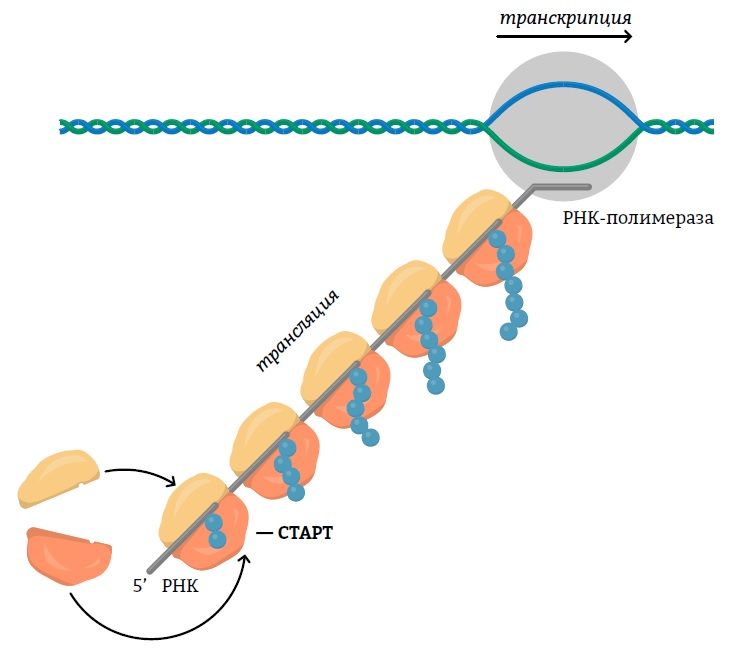
У прокариот процессы транскрипции и трансляции проходят одновременно, поэтому матричная РНК имеет только первичную структуру. Так же как и у эукариот, она представлена линейной последовательностью рибонуклеотидов, которая содержит информационные и некодирующие участки.
Большинство мРНК бактерий и архей полицистронны (содержат несколько кодирующих областей), что обусловлено особенностью организации прокариотического генома, который имеет оперонную структуру. Это означает, что в одном транскриптоне ДНК закодирована информация о нескольких белках, которая впоследствии переносится на РНК. Небольшая часть информационной РНК является моноцистронной.
Нетранслируемые области бактериальной мРНК представлены:
- лидерной последовательностью (расположена на 5`-конце);
- трейлерной (или концевой) последовательностью (находится на 3`-конце);
- нетранслируемыми межцистронными областями (спейсерами)- находятся между кодирующими участками полицистронной РНК.
Длина межцистронных последовательностей может состоять от 1-2 до 30 нуклеотидов.

Эукариотическая мРНК
Эукариотическая мРНК всегда моноцистронна и содержит более сложный набор некодирующих областей, которые включают:
- кэп;
- 5`-нетранслируемую область (5`НТО);
- 3`-нетранслируемую область (3`НТО);
- полиадениловый хвост.
Обобщенную структуру матричной РНК у эукариот можно представить в виде схемы со следующей последовательностью элементов: кэп, 5`-НТО, AUG, транслируемая область, стоп-кодон, 3`НТО, поли-А-хвост.

У эукариот процессы транскрипции и трансляции разобщены и во времени и в пространстве. Кэп и полиадениловый хвост матричная РНК приобретает во время созревания, которое называется процессингом, а затем транспортируется из ядра в цитоплазму, где сосредоточены рибосомы. В ходе процессинга также вырезаются интроны, которые переносятся на РНК с эукариотического генома.
Где синтезируются рибонуклеиновые кислоты
Все виды РНК синтезируются специальными ферментами (РНК-полимеразами) на основе ДНК. Соответственно, локализация этого процесса в прокариотической и эукариотической клетках отличается.
У эукариот транскрипция осуществляется внутри ядра, в котором сосредоточена ДНК в виде хроматина. При этом вначале синтезируется пре-мРНК, которая претерпевает ряд модификаций и только после этого транспортируется в цитоплазму.
У прокариот местом, где синтезируются рибонуклеиновые кислоты, служит область цитоплазмы, граничащая с нуклеоидом. РНК-синтезирующие ферменты взаимодействуют с деспирализованными петлями бактериального хроматина.
Механизм транскрипции
Синтез матричной РНК основан на принципе комплементарности нуклеиновых кислот и осуществляется РНК-полимеразами, которые катализируют замыкание фосфодиэфирной связи между рибонуклеозидтрифосфатами.
У прокариот мРНК синтезируется тем же ферментом, что и другие виды рибонуклеотидов, а у эукариот — РНК-полимеразой II.

Транскрипция включает 3 стадии: инициацию, элонгацию и терминацию. На пером этапе полимераза присоединяется к промотору — специализированному участку, предваряющему кодирующую последовательность. На стадии элонгации фермент наращивает цепь РНК, присоединяя к цепи нуклеотиды, комплементарно взаимодействующие с матричной цепью ДНК.